Main Model
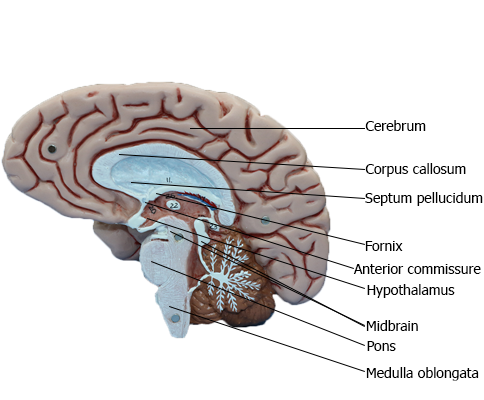
BRAIN : Medial view of right side of brain
The Telencephalon
The telencephalon is the largest part of the human brain, constituting about 85% of total brain weight, and is that portion in
which all modalities are represented. Various sensory inputs (such
as vision and hearing) are localized in some areas, whereas motor
functions are represented in other regions and are modulated
by subcortical nuclei. The telencephalon contains circuits that
interrelate regions that have specific functions, such as motor
or visual, with other regions called association areas. Seeing a
familiar image may precipitate a cascade of neural events having
olfactory, emotional, sensory, and motor components. Damage
to association areas results in complex neurologic deficits. The
patient may not be blind or paralyzed but may be unable to recognize sensory input (agnosia), express ideas or thoughts (aphasia), or perform complex goal-directed movements (apraxia).
Overview
The telencephalon consists of large hemispheres separated from
each other by a deep longitudinal cerebral fissure. Each hemisphere has an outer surface, the cerebral cortex, which is composed of layers of cells. The cortex is thrown into elevations
called gyri (singular, gyrus) that are separated by grooves called
sulci (singular, sulcus). Internal to the cortex are large amounts
of subcortical white matter along with aggregates of gray matter
that form the basal nuclei and the amygdala. Although not parts
of either the telencephalon or the basal nuclei, the subthalamic nucleus (of the diencephalon) and the substantia nigra (of the
mesencephalon) have important connections that functionally
link them with the basal nuclei.
Information passing into or out of the cerebral cortex must
traverse the subcortical white matter. The myelinated fibers
forming the white matter are organized into (1) association bundles that connect adjacent or distant gyri in one hemisphere; (2) commissural bundles that connect the hemispheres, the largest of these being the corpus callosum; and (3) the internal capsule. The internal capsule contains axons projecting to numerous
downstream nuclei (corticofugal fibers) and axons conveying
information to the cerebral cortex (corticopetal fibers). The
terms corticofugal and corticopetal are umbrella terms that
include all efferent and all afferent fibers, respectively, of the
cerebral cortex.
The hippocampal complex and the amygdala are located in
the walls of the temporal horn of the lateral ventricle. The axons
of cells in these structures coalesce to form the fornix, stria terminalis, and amygdalofugal pathway.
Development
Enlargements of the prosencephalon, the telencephalic (cerebral) vesicles, appear at about 5 weeks of gestation. As the cerebral vesicles enlarge in all directions, they pull along portions of the neural canal that will form the cavities of the telencephalon,
the lateral ventricles. The primitive lateral ventricles extend into frontal, parietal, temporal, and occipital areas as
they develop and form that portion of the ventricle found in each
of these lobes in the adult. The interventricular foramina, which
connect each lateral ventricle to the midline third ventricle (cavity of the diencephalon), are initially large but become smaller as
development progresses. In the adult brain, each interventricular
foramen is bordered rostromedially by the column of the fornix
and caudolaterally by the anterior tubercle of the dorsal thalamus.
Cells forming the corpus striatum appear in the floor of the
developing lateral ventricle at the time when primordial cell
groups in the wall of the third ventricle are giving rise to diencephalic structures. As development progresses,
the corpus striatum is bisected by axons growing to and from the
cerebral cortex. These axons form the internal capsule of the
adult and divide the corpus striatum into a medially located caudate nucleus and a laterally located putamen. As the diencephalon enlarges, it gives rise to the thalamus and hypothalamus and
to cells that migrate across the developing internal capsule to
assume a position medial to the putamen. These
cells become the globus pallidus of the adult and, in combination
with the putamen, form the lenticular nucleus.
The initial development of the major commissural bundles and
of the hippocampus takes place along the medial aspect of the
hemisphere. In the adult brain, there are three
major interhemispheric commissures: the anterior commissure,
the hippocampal commissure, and the corpus callosum. The first
of these to appear, the anterior commissure, arises within the lamina terminalis, a membrane-like structure that extends from the anterior commissure anteriorly (ventrally) to the rostral edge
of the optic chiasm. The second to form, the hippocampal commissure, develops along with the hippocampal primordium. As growth occurs, the hippocampus, which originates
in the posteromedial part of the hemisphere, is displaced into the
temporal lobe, where it assumes a position characteristic of the
adult. In the process, fibers from one side cross to
the other side, as the hippocampal commissure, just inferior to the area that will be occupied by the corpus callosum. The third
commissure to develop, the corpus callosum, originates from the
area of the lamina terminalis as a structure initially composed
of astrocytic processes. Axons from developing neurons in each
hemisphere traverse this glial structure to access the contralateral
side. As this takes place, the corpus callosum enlarges in a caudal direction to form the prominent structure found in the adult.
Developmental Defects
There are numerous developmental events that may cause
defects in the configuration of the telencephalon. One of the developmental failures that will
result in aberrant development of the telencephalon is the
improper migration of maturing neurons on radial glia. This
failure results in structural and in some cases corresponding
functional defects in the arrangement of the cerebral cortex.
Some examples include lissencephaly (a lack of gyri and sulci,
a smooth brain), pachygyria (abnormally large gyri that are
few in number), and microgyria (abnormally small gyri that
are greater in number).
Holoprosencephaly is a preneurulation defect that is represented by three general forms. Alobar holoprosencephaly, the
most severe form, consists of a midline ventricle, no hemispheres
or corpus callosum, and severe retardation. Semilobar holoprosencephaly consists of a partial formation of lobes with the ventricles formed; the frontal lobes may be fused, and the occipital lobes may be separated by an incomplete longitudinal fissure.
Although the ventricles are formed, midline structures such as
the septum pellucidum are missing. In lobar holoprosencephaly,
the least severe form, the longitudinal fissure is largely complete, hemispheres exist, generally normal patterns of sulci and gyri are seen, but there is a fusion of the hemispheres at the frontal pole
or at the orbital surface of the frontal lobe.
Anencephaly is a severe developmental failure in which the
telencephalon and the surrounding skull are largely absent. This
defect is catastrophic and not compatible with life. Anencephaly
is generally associated with a failure of the anterior neuropore to
close. The lamina terminalis represents the adult position of the
anterior neuropore.
Failure of the corpus callosum to develop (agenesis of the corpus callosum) may be accompanied by an absence of the anterior
and hippocampal commissures. Although some patients with this condition may experience focal seizures and
have mental retardation, others live for many years with few
or no obvious neurologic deficits. These individuals frequently have developmental abnormalities in other parts of the nervous
system.
---------------------------------------------------------------------------------------------------------------------------------------------------------------
Hippocampus and Amygdala
The hippocampal formation and the amygdaloid complex are
located in the temporal lobe. The former lies in the inferomedial
floor of the temporal horn of the lateral ventricle and the latter in the rostral end of this space. Through a variety of pathways,
these structures interconnect with numerous telencephalic and
diencephalic centers.
Developmentally, the hippocampus is formed by an invagination of primitive cortex to form the curved, multilayered
structure characteristic of the adult brain. The hippocampal formation is found internal to the parahippocampal gyrus and is composed of the subiculum, the hippocampus
proper (also called Ammon horn), and the dentate gyrus. The
cortex of the parahippocampal gyrus is continuous with the
subiculum, which in turn is continuous with the hippocampus
proper. The dentate gyrus forms a reverse loop adjacent to the
hippocampus and, in doing so, presents a serrated surface that
is medially exposed to the subarachnoid space.
Axons of hippocampal neurons converge to form a prominent
bundle that arches around caudal, superior, and rostral aspects of
the thalamus. This bundle, the fornix, is a major efferent path of
the hippocampal formation. It is composed of a flattened caudal part, the crus; a compact superior
portion, the body; and a part that arches around the rostral part
of the thalamus and passes through the hypothalamus to terminate in the mammillary body - this is the column of the fornix.
Located along the edge of the dentate gyrus and continuing on
the lateral edge of the crus and body of the fornix is a thin fringe
of fibers called the fimbria.
The amygdaloid nuclear complex (commonly called the
amygdala) is located internal to the cortex of the uncus. It is composed of several cell groups,
including caudomedial, basolateral, and central subdivisions.
Two major efferent bundles are related to the amygdala. First,
the stria terminalis follows a looping trajectory that shadows, in
a reverse direction, the orientation of the caudate nucleus. In the temporal horn, the stria terminalis is located
just medial to the tail of the caudate nucleus. As the stria terminalis arches superiorly and rostrally, it assumes a position in
the shallow groove between the caudate nucleus and the dorsal
thalamus, where it is accompanied by the terminal vein (superior thalamostriate vein). At about the level of
the interventricular foramen, the fibers of the stria terminalis
fan out to enter and terminate in the hypothalamus, the septal
area, and the neostriatum.
The second major efferent bundle of the amygdala is the diffusely arranged ventral amygdalofugal pathway. These fibers
leave the amygdaloid complex, pass medially through the substantia innominata, and continue medially to enter hypothalamic and septal nuclei or turn caudally and distribute to the
brainstem.
Cell groups located internal to the subcallosal area collectively
form the septal nuclei. Consequently, the subcallosal area, together with a small strip of cortex located adjacent to the lamina terminalis, the paraterminal gyrus, is commonly called the
septal area. The septal nuclei are medially adjacent to the nucleus
accumbens and continuous with sheets of neuronal cell bodies that
extend into the septum pellucidum. The latter structure extends,
in general, from the fornix to the inner surface of the corpus callosum. It forms the medial wall of the anterior horns and a small
part of the bodies of the lateral ventricles.
In general, the septal nuclei have complex interconnections with
hippocampal, amygdaloid, and other limbic structures.
---------------------------------------------------------------------------------------------------------------------------------------------------------------
An Overview of the Brainstem
The term brainstem (sometimes written brain stem) can mean
either the portion of the brain that consists of the medulla oblongata, pons, and midbrain or the portion that consists of these structures plus the diencephalon. Following the former
convention, for our purposes, the brainstem consists
of the rhombencephalon (excluding the cerebellum) and the mesencephalon. These regions of the brainstem share a basic
organization.
Basic Divisions of the Brainstem
Medulla Oblongata
At about the level of the foramen magnum, the spinal cord
merges into the most caudal portion of the brain, the medulla
oblongata or myelencephalon, commonly called the medulla.
The foramen magnum marks the approximate location of the
motor (pyramidal) decussation of the medulla. The
medulla is slightly cone shaped and enlarges in diameter as it
extends rostrally from the medulla-spinal cord junction toward
the pons-medulla junction. On the posterior (dorsal) aspect of
the medulla-pons continuum, this junction is represented by the
caudal edge of the middle and inferior cerebellar peduncles,
whereas anteriorly (ventrally) this border is formed by the caudal
edge of the basilar pons.
The cranial nerves associated with the medulla include the
hypoglossal (XII, motor) and parts of the accessory (XI, motor),
vagus (X, mixed), and glossopharyngeal (IX, mixed) nerves. The nuclei of the hypoglossal, vagal, and glossopharyngeal nerves as well as portions of the nuclei of the trigeminal
nerve are located in the medulla.
The abducens (VI, motor), facial (VII, mixed), and vestibulocochlear (VIII, sensory) nerves are frequently called the cranial
nerves of the pons-medulla junction because they exit the brainstem at this particular location.
Although the medulla does not have regions that are specifically regarded as tegmental or basilar (as is the case for the pons
and midbrain), it does have regions that function in the same way
and are rostrally continuous with these respective regions of the
pons. For example, the central regions of the medulla
contain the cranial nerve nuclei affiliated with the medulla. This
medullary area is rostrally continuous with the pontine tegmentum, which contains the cranial nerve nuclei associated with the
pons. In similar manner, the pyramids of the medulla (containing
corticospinal fibers) are located on the anterior ("basal") aspect
of the medulla and are rostrally continuous into the basilar pons.
Pons
The pons (the anterior part of the metencephalon) extends from
the pons-medulla junction to an imaginary line drawn from the
exit of the trochlear nerve posteriorly to the rostral edge of the
basilar pons anteriorly. What we commonly call
the pons is actually composed of two portions, the pontine tegmentum (located internally) and the basilar pons.
The basilar pons is bulbous and quite characteristic of the anterior
aspect of the pons. The pontine tegmentum contains portions of
the trigeminal nuclei and the vestibular nuclei and, just rostral
to the pons-medulla junction, the facial motor nucleus, superior salivatory nucleus, and abducens nucleus. The trigeminal
nerve (V, mixed) emerges from the lateral aspect of the pons, and
abducens (VI), facial (VII), and vestibulocochlear (VIII) nerves
exit at the pons-medulla junction.
The cerebellum, although part of the metencephalon, is not
part of the brainstem. It is joined to the brainstem by three large,
paired bundles of fibers called the cerebellar peduncles. These are
the inferior cerebellar peduncle, the middle cerebellar peduncle (or brachium pontis), and the superior cerebellar peduncle
(or brachium conjunctivum), connecting the cerebellum to the medulla oblongata, basilar pons, and midbrain, respectively.
Midbrain
The midbrain (mesencephalon) extends rostrally from the pons-midbrain junction to join the diencephalon (thalamus); this
interface is usually described as a line drawn from the posterior commissure posteriorly to the caudal edge of the mammillary
bodies anteriorly. The oculomotor nerve (III, motor)
exits the anterior aspect of the midbrain, whereas the trochlear nerve (IV, motor) exits its posterior aspect. The
exit of the trochlear nerve is regarded as the pontomesencephalic
junction on the posterior aspect of the brainstem; along with its decussating fibers, it composes the isthmus rhombencephali
(the transition from pons to midbrain).
The posterior aspect of the midbrain is characterized by the
superior and inferior colliculi and the anterior aspect by the crus
cerebri and interpeduncular fossa.
Tegmental and Basilar Areas
The central core of the midbrain and the pons is called the tegmentum, and their anterior (ventral) parts are the basilar areas.
These regions are continuous with each other and with comparable areas of the medulla. The tegmentum of the pons and midbrain and the contiguous central portion
of the medulla contain ascending and descending tracts, many
relay nuclei, and the nuclei of cranial nerves III to XII.
The basilar part of each brainstem division is anterior to the
tegmentum (of the midbrain and pons) and to the central portion of the medulla. Consequently, these basilar structures also form a rostrocaudal continuum. Basilar structures of
the brainstem include the descending fibers of the crus cerebri (midbrain), basilar pons, and pyramid (medulla) and specific populations of neurons in the midbrain and pons that originate
from the alar plate of the embryonic brain.