Main Model
Anterior : Nucleus
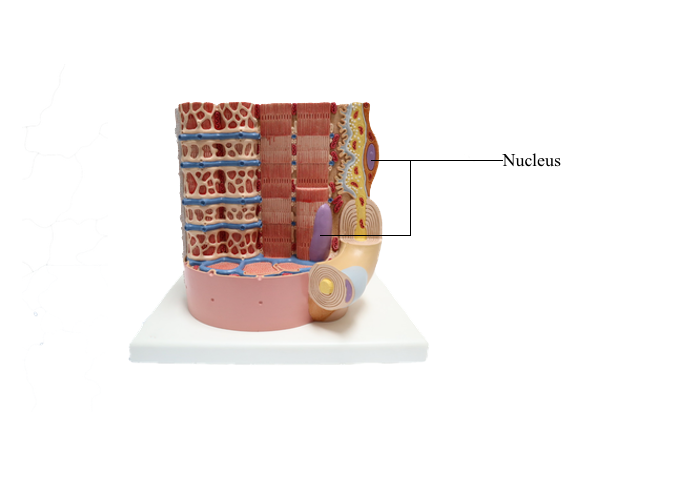
Cell Nucleus
Nuclear Envelope and Nuclear Pore Complex
The cell nucleus consists of three major components:
1. The nuclear envelope.
2. Chromatin.
3. The nucleolus.
The nuclear envelope consists of two concentric
membranes separated by a perinuclear space. The inner nuclear membrane is associated with the nuclear
lamina, chromatin, and ribonucleoproteins. The outer nuclear membrane is continuous
with the membranes of the endoplasmic reticulum
and can be associated with ribosomes.
The nuclear pore complex has a tripartite structure, composed of a central cylindrical body placed
between inner and outer octagonal rings, each consisting of eight protein particles. The central cylinder
consists of a central plug and eight radiating spokes. The exact role of individual nuclear
pore complex proteins in nucleocytoplasmic trafficking is unclear.
Nuclear pore complexes embedded in the nuclear
envelope establish bidirectional communication gates
for the trafficking of macromolecules between the
cytoplasm and the nucleus. Small molecules (less
than 40 to 60 kd) can diffuse passively through the
nuclear pore complex. Proteins of any size, containing
a nuclear localization amino acid sequence (NLS,
Pro-Lys-Lys-Lys-Arg-Lys-Val), can be imported
into the nucleus, however, by an energy-dependent mechanism (requiring ATP and GTP).
Nucleocytoplasmic Transport: Ran-GTPase
Protein nuclear import/export is controlled by Ran
(for Ras-like nuclear GTPase), a small GTPase of
the Ras superfamily that dictates the directionality
of nucleocytoplasmic transport.
Ran shuttles across the nuclear pores and accumulates inside the nucleus by an active transport
mechanism.
1. In the nucleus, a high concentration of Ran-GTP is achieved by RCC1, a GDP-GTP exchanger protein bound to chromatin. Ran-GTP determines
the dissociation of imported proteins containing NLS
by binding to importin beta, the transporter receptor
protein.
2. In the opposite direction, from the nucleus to
the cytoplasm, binding of Ran-GTP to the carrier
protein exportin/Crm1 facilitates the assembly of
complexes containing proteins with nuclear export
sequence (NES).
3. In the cytoplasm, Ran-GTP is converted to
Ran-GDP by Ran-GTPase, which is activated by two cooperating proteins: Ran-GAP (Ran-GTPase-activating protein) and RanBP (Ran-GTP binding
protein). Consequently, the exported protein is dissociated from its transporter receptor protein exportin/
Crm1 and Ran-GTP. Importin and exportins are
recycled by transport back across the nuclear pore
complex.
Chromatin
Chromatin is defined as particles or "beads" (called
nucleosomes) on a double-stranded DNA string. Each nucleosome consists of a histone
octamer core and about two turns of DNA wound around the histone core. The histone octamer contains two molecules each of H2A, H2B, H3, and H4
histones. H1 histone cross-links the DNA molecule
wrapped around the octamer.
Chromatin is packed in separate chromosomes that
can be visualized during mitosis (or meiosis). During
interphase (phases G1, S, and G2 of the cell cycle),
individual chromosomes cannot be identified as such,
but are present in a diffuse or noncondensed state.
Diffuse chromatin, called euchromatin ("good
chromatin"), is transcriptionally (RNA synthesis) active and represents about 10% of total chromatin. Euchromatin is the site of synthesis on nonribosomal
RNAs, including mRNA and transfer RNA (tRNA)
precursors.
Condensed chromatin, called heterochromatin
("different chromatin"), is transcriptionally inactive and represents about 90% of total chromatin.
Dosage Compensation: X Chromosome Inactivation
X chromosome inactivation, known as dosage
compensation, starts early in embryonic stem cell
differentiation and is characterized by four features:
1. All but one of the X chromosomes undergoes
inactivation.
2. The choice of the inactivated X chromosome
is random. Either the paternal or the maternal X
chromosome is inactivated.
3. The inactivation processes is heritable through
subsequent rounds of cell division. The choice remains nonrandom for all subsequent cell descendants.
4. Both X chromosomes in oocytes remain active.
The transcriptional inactivation of one of the two X
chromosomes is observed in the trophoblast on day
12 after fertilization and on day 16 in the embryo.
In humans, the inactivated X chromosome is
recognized by the presence of the Barr body, a heterochromatin mass observed adjacent to the
nuclear envelope or in the form of a drumstick in
polymorphonuclear leukocytes. If
a cell has more than two X chromosomes, the extra
X chromosomes are inactivated, and more than one
Barr body is visualized.
The concept of dosage compensation is relevant
to the understanding of tumor-suppressor inactivation and oncogene inactivation when a single active
copy of an X-linked genes is affected. Some genes
located on the inactivated X chromosome escape
inactivation in normal cells and several of these genes,
most of which encode growth factors, are implicated
in human cancer. For example, the gene encoding gastrin-releasing peptide receptor is associated with
an increased risk in lung cancer in women.
Nucleolus
The nucleolus is the site of synthesis and processing of ribosomal RNA (rRNA) and assembly of ribosomal subunits. The rRNA genes are arranged
in an array of multiple copies transcribed by RNA
polymerase I.
The nucleolus houses several proteins, including
fibrillarin and nucleolin, required for pre-rRNA processing. In addition, the nucleolus contains nucleostemin, a protein unrelated to ribosomal biogenesis.
Nucleolin and nucleostemin are shuttling proteins;
they relocalize from the nucleolus to the nucleoplasm
where they interact with protein p53, a protector
of DNA damage by preventing DNA replication in
response to genomic stress.
Essentially, the nucleolus is a multifunctional
nuclear structure consisting of stable proteins involved in ribosomal synthesis and molecules shuttling
between the nucleolus and nucleoplasm to fulfill
non-nucleolar functions.
Structurally, the nucleolus consists of three major
components:
1. A fibrillar center (corresponding to chromatin
containing repeated rRNA genes and the presence of
RNA polymerase I and signal recognition particle
[SRP] RNA).
2. A dense fibrillar component (where nascent
rRNA is present and undergoing some of its processing). Fibrillarin and nucleolin are found in the
fibrillar dense component.
3. A granular component (where the assembly of
ribosomal subunits, containing 18S rRNA [small
subunit] and 28S rRNA [large subunit], is completed). Nucleostemin, a protein unrelated to ribosomal
biogenesis, coexists with the granular components.
Nucleoli are typically surrounded by a shell of
heterochromatin, mostly from centromeric and pericentromeric chromosomal regions.
The nucleolus dissociates during mitosis, then reappears at the beginning of the G1 phase. More than
one nucleolar mass, each representing the product of
a chromosome with a nucleolar organizing region
(NOR), can be observed in the nucleus. In some
cells with an extended interphase, such as neurons,
a single large nucleolus is organized by the fusion of several nucleolar masses.
The active process of rRNA synthesis can be visualized at the electron microscopic level by spreading the contents of nuclei of cells with
hundreds of nucleoli (e.g., amphibian oocytes). rRNA
genes can be seen as repeating gene units along the
chromatin axis, like "Christmas trees," pointing in
the same direction and separated by nontranscribed
spacers. The entire rRNA gene region is covered by
more than 100 RNA polymerase I molecules synthesizing an equivalent number of fibrils, each with
a terminal granule.
Each fibril represents an rRNA precursor (45S)
ribonucleoprotein molecule oriented perpendicularly to the chromatin axis similar to the branches of
a tree. The 45S rRNA precursor is detached from
the chromatin axis and cleaved into 28S, 18S, and
5.8S rRNAs.
The 18S rRNA and associated proteins form the
small ribosomal subunit. The 28S and 5.8S, together
with 5S rRNA made outside the nucleolus, and associated proteins form the large ribosomal subunit.
The mRNA precursor is transcribed by RNA polymerase II, and the tRNA precursor is transcribed by
RNA polymerase III.
Localization of Nucleic Acids
Cytochemistry and autoradiography provide information about the cellular distribution
and synthesis of nucleic acids. The Feulgen reaction
is specific for the localization of DNA. Basic dyes, such as toluidine blue, stain DNA and
RNA. Pretreatment with deoxyribonuclease (DNAse) and ribonuclease (RNAse) defines
the distribution sites of DNA and RNA by selective removal of one of the nucleic acids.
Autoradiography and radiolabeled precursors for
one of the nucleic acids can determine the timing of
their synthesis. In this technique, a radioactive precursor of DNA ([3H]thymidine) or RNA ([3H]uridine)
is exposed to living cells. As a result of exposure to the
radiolabel, any synthesized DNA or RNA contains
the precursor. The radioactivity is detected by coating
the cells with a thin layer of a photographic emulsion.
Silver-containing crystals of the emulsion are exposed
to structures of the cell containing radioactive DNA
or RNA. After development of the emulsion, silver
grains indicate the location of the labeled structures.
This approach has been used extensively for determining the duration of several phases of the cell cycle.